Eucoilinae
Numerous images are available at the MorphBank site, http://www.morphbank.net, relative to species and taxa discussed here and in the other eucoiline pages on the Paroffit site. See the following pages for additional information Aganaspis, Dicerataspis, Ealata, and Odontosema.
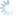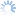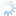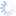
Leptopilina is the most commonly encountered genus of parasitoids reared from fruit-inhabiting Drosophilidae, and is thus included in this key for comparison with those more frequently associated with Tephritidae.
1. Surface of the fore wing bare or nearly so (Fig. 1: Odontosema and Figs. 2,3: Lopheucoila) (go to 2)
1a. Surface of fore wings distinctly setose (Figs. 4, 5) (go to 3)
2. Scutellar disc, in dorsal view, with two tooth-like lobes posteriorly (Fig. 6). Scutellar plate flat (Fig. 7). Odontosema
2a. Scutellar disc, in dorsal view, with four tooth-like lobes (Fig. 8). Scutellar plate bearing a small, spinose bump on its dorsal surface (Fig. 9). Lopheucoila
3. Scutellar disc with tooth-like lobes posteriorly (Fig. 10). Dicerataspis
3a. Scutellar disc truncate or more or less evenly rounded, never with tooth-like lobes posteriorly (Fig. 11) (go to 4).
4. Scutellar plate bearing a small, spinose bump on its dorsal surface (Figs. 12, 13), similar to that found in Lopheucoila. Originally described from Afrotropical Region, but subsequently recorded from Taiwan. Ealata
4a. Scutellar plate flat, never bearing spinose bump on dorsal surface (Figure 11). (go to 5)
5. Pronotal plate protrudes above the anterior margin of the mesoscutum. Scutellar plate generally broader and flatter. Fourth antennal segment in males is equal to or shorter than third, and not bent (Fig. 14). Aganaspis
5a. Pronotal plate does not protrude above anterior margin of the mesoscutum. Scutellar plate often narrower and more convex. Fourth antennal segment of males usually longer than the 3rd, and usually bent. Leptopilina














Additionally, there are published records for Dicerataspis and Lopheucoila that strongly suggest that members of these genera at least occasionally attack Tephritidae even though they normally attack hosts in other families. At least one species of Ealata has been also reared on several occasions from fruit infested with tephritids, but hosts are as yet unconfirmed. Finally, there are records of Rhoptomeris and Trybliographa attacking tephritids in the New World, but these are likely misidentifications of some of the more difficult and poorly defined New World genera of eucoilines. A revision of the New World genera will be necessary before these host relationships can be clarified. Wharton et al. (1998) provide a detailed summary of published records referring to species of Rhoptromeris and Trybliographa that have been associated with tephritids.
Many of the eucoilines reared from fruits are actually parasitoids of Drosophilidae, Lonchaeidae, Phoridae or other flies associated with ripe and decomposing fruit. Members of the genus Leptopilina, for example, are well-known as parasitoids of Drosophilidae and are among the mostly commonly encountered eucoilines reared from fruit samples in the Afrotropics. Though Drosophilidae tend to colonize fruit after tephritids have departed to pupate in the soil, there is potential for overlap. For example, fruits that have fallen from a tree may have more rapidly decaying portions inhabited by Drosophilidae at the same time as more sound portions still inhabited by tephritid larvae. Since fruit is often collected and reared in bulk, it is easy to obtain parasitoids of both Drosophilidae and Tephritidae from the same sample. Thus, in order to assess host specificity accurately, it is essential to verify all records by isolating individual puparia or exposing known hosts to ovipositing females. For many of the older records (including label data from unpublished studies), this was not done and host associations must therefore be viewed with caution. It is likely that the tephritid host records for Rhoptromeris haywardi (Blanchard) and Trybliographa brasiliensis (von Ihering) (e.g. Von Ihering 1905, 1912) actually refer to drosophilids for these reasons. Puparia of most fruit-infesting tephritids do not have the posterior spiracles protruding from the surface and this will help in distinguishing them from many of the other Diptera noted here.
Since most fruit-infesting tephritids are considerably larger than fruit-infesting drosophilids, and their eucoiline parasitoids are solitary, it is therefore often possible to distinguish drosophilid parasitoids from tephritid parasitoids simply by differences in their size. Some tephritids are quite small, however, making such associations somewhat risky in the absence of isolated puparia.
